Sekretion
Als Sekretion (lateinisch secretio „Absonderung“; Verb dazu ist sezernieren, von lateinisch secernere „absondern“) oder Absonderung wird die Abgabe von flüssigen Stoffen, die (im Gegensatz zum Exkret bei der Exkretion) eine bestimmte Funktion erfüllen, durch einzelne Zellen oder Drüsen bezeichnet. Das Sekret (lat. Secretum) wird unwillkürlich abgeschieden; die Regelung geschieht durch das vegetative Nervensystem sowie durch Rückkopplungsmechanismen, an denen teilweise Hormone beteiligt sind.
Einteilung
Nach Richtung der Sekretion
Werden die Stoffe an innere oder äußere Körperoberflächen abgegeben, spricht man von exokriner Sekretion. Werden sie dagegen in den Extrazellularraum abgegeben, spricht man von innerer Sekretion; gelangt das Sekret von dort aus in den Blutkreislauf, spricht man von endokriner Sekretion oder Inkretion und das Drüsenprodukt wird auch als Inkret bezeichnet. Den Begriff „innere Sekretion“ prägte 1855 Claude Bernard. Die bereits von Georg Ernst Stahl im 18. Jahrhundert geförderte und 1849 von Arnold Adolf Berthold experimentell nahegelegte Lehre von der inneren Sekretion[1] wird als Endokrinologie bezeichnet.
Andere Formen der inneren Sekretion sind die autokrine Sekretion (Wirkung des Sekrets auf die absondernde Zelle selbst) und die parakrine Sekretion (Wirkung des Sekrets auf die unmittelbare Umgebung).
Nach Sekretionsmechanismus
- Ekkrine Sekretion: Transport von im Zytosol gelösten kleinen Ionen oder Molekülen über die Zellmembran mittels Transportproteinen. Wenn osmotisch aktive Teilchen transportiert werden, folgt Wasser nach, worin oft der eigentliche Zweck dieser Sekretion besteht. Beispiel: Schweißdrüsen des Menschen.
- Merokrine Sekretion: Sekretion per Exozytose: Die sezernierten Stoffe befinden sich in der Zelle in Vesikeln, deren Inhalt durch Verschmelzung mit der Zellmembran nach außen freigesetzt wird. Dies ist der klassische Weg für die Sekretion von Proteinen. Beispiele: Ohrspeicheldrüse, exokrine Bauchspeicheldrüse, Milchdrüse (Proteine und Milchzucker). Exokrine merokrine Drüsen betreiben zusätzlich ekkrine Sekretion, um die Flüssigkeit zu bilden, in der die Proteine gelöst sind.
- Apokrine Sekretion: Sekretvesikel werden mit umgebendem apikalem Zytoplasma durch einen Teil der Zellmembran von der Drüsenzelle abgeschnürt. Beispiele: Duftdrüsen des Menschen, Prostata, Bläschendrüse, Milchdrüse (Fetttröpfchen).
- Holokrine Sekretion: Die ganze Zelle wird zur Sekretbildung abgegeben und geht zugrunde. Das Sekret füllt die Zelle aus, der Kern wird pyknotisch und schließlich zerfällt die Zelle. Beispiele: Talgdrüsen, Kropfmilch.
Nach Funktion des Sekrets
Ein Sekret kann unterschiedliche Aufgaben erfüllen:
- Feuchthalten der Schleimhäute und Bindung von Krankheitserregern (zum Beispiel Nasensekret)
- Verdauung innerhalb des Körpers (zum Beispiel Speichel, Magensaft, Galle)
- Verdauung außerhalb des Körpers (zum Beispiel bei Spinnen)
- Ernährung des Nachwuchses (Milchdrüse der Säugetiere)
- Thermoregulation (zum Beispiel Schweiß)
- Einfettung von Haut und Haaren (Talgdrüsen)
- Absonderung von Duftmarken (Duftdrüsen)
- Zusetzung von Nährmedien (zum Beispiel Akzessorische Geschlechtsdrüsen)
- Offenhalten von Stich- oder Bisswunden (zum Beispiel bei Stechmücken, Blutegeln und Vampirfledermäusen)
- Abwehr von Fraßfeinden durch Gifte oder stinkende Sekrete (Wehrsekret, Reflexbluten)
- Vergiftung von Beutetieren (zum Beispiel Schlangengift)
Dabei kann ein Sekret gleichzeitig mehrere dieser Funktionen erfüllen: Die Galle dient beispielsweise sowohl der Ausscheidung als auch der Verdauung.
Zelluläre Sekretion
Auf zellulärer Ebene wird auch die Ausschleusung eines einzelnen Proteins als Sekretion bezeichnet. Diese Proteine (zum Beispiel Immunglobuline (Antikörper) oder Bestandteile der extrazellulären Matrix) heißen dann auch sekretorische Proteine.
Ferner unterscheidet man die konstitutive Sekretion von der regulierten Sekretion (siehe auch Exozytose):
- Die konstitutive Sekretion betrifft Proteine ohne besondere Signale (Signaltransduktion). Sie verbleiben in sogenannten default-Vesikeln, die mit der Membran verschmelzen und das Protein sezernieren.
- Bei der regulierten Sekretion werden Proteine in spezialisierte Vesikel verpackt, nachdem sie an einen spezifischen Rezeptor gebunden haben. In diesen Vesikeln können sie noch einmal modifiziert oder gespeichert werden, bis ein Reiz die Zelle zur Sekretion stimuliert. Hierbei haben die Proteine Secretogranin II und Chromogranin B wahrscheinlich eine bestimmte Funktion, denn sie sorgen für das Aggregieren der auszuschüttenden Proteine. Dieser Prozess ist jedoch noch nicht sicher erforscht.
Eukaryotische Proteinsekretion
Der Sekretionsweg ist grundsätzlich in allen eukaryotischen Zellen gleich: Vesikel bewegen die produzierten Proteine aus dem Endoplasmatischen Retikulum (ER) über den Golgi-Apparat zur Plasmamembran. Nur die Menge der produzierten sekretorischen Proteine variiert. Zum Beispiel werden in Pflanzenzellen Produkte wie Kollagen sezerniert, während im menschlichen Verdauungssystem einige Enzyme produziert werden.[2]
Schritte des sekretorischen Weges
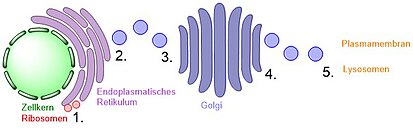
Erstens synthetisieren Ribosomen in der Membran des Endoplasmatischen Retikulums Proteine. Dies wird als Translation bezeichnet. Diese Proteine werden entweder cotranslational in die ER-Membran eingebunden (integrale Proteine) oder treten in das ER-Lumen ein, wo ihr Signalpeptid abgespalten wird. Dort werden die Proteine gefaltet und es kann eine N-Glykosylierung stattfinden.[3]
Zweitens werden Proteine in Transportvesikel verpackt. Diese Transportvesikel lösen sich von dem Spenderorganell ab und verschmelzen mit der Zielmembran, um ihre Fracht (= Proteine) freizusetzen.
Der dritte Schritt ist die Verschmelzung der Transportvesikel entweder mit der cis-Seite des Golgi-Apparates oder mit einem weiteren Vesikel zur Bildung eines neuen cis-Golgis. Vom cis-Golgi gehen auch einige Vesikel zurück zum ER.
Danach bewegen sich die Proteine innerhalb des Golgi von der cis- zur trans-Seite, während der Golgi-Apparat O-Glykosylierungen einfügt oder die N-Glykosylierungen modifiziert. Wenn das Protein zum Lysosomen transportiert werden soll, erhält es außerdem eine Mannose-6-phosphat-Markierung.
Abschließend werden die modifizierten Proteine entweder vom trans-Golgi zur Plasmamembran transportiert[2] oder zum Lysosomen. Die Anfangs als integrale Proteine synthetisierten Proteine befinden sich auch nun immer noch in der Membran.
Verschiedene Transportvesikel für verschiedene Routen
Im Allgemeinen haben alle Zellkompartimente unterschiedliche molekulare Marker auf ihrer Membran. Ähnlich haben Transportvesikel unterschiedliche Mäntel, die unterschiedliche Ladung auswählen. Untereinheiten in den Mänteln führen sie zu verschiedenen Zellkompartimenten. Die drei Hauptsorten beschichteter Vesikel sind Clathrin-Vesikel, COPI-Vesikel und COPII-Vesikel. COPII-Vesikel knospen vom Endoplasmatischen Retikulum ab. COPI-Vesikel transportieren Proteine innerhalb des Golgi von Cisternae zu Cisternae. Clathrin-Vesikel transportieren Proteine vom trans-Golgi entweder zu Endosomen oder direkt zur Plasmamembran.[3]
Bakterielle Sekretion
Siehe auch
- Tubulus #Funktion der Tubuli der Nieren
Literatur
- Rudolf Heidenhain: Physiologie der Absonderungsvorgänge. In: Hermanns Handbuch der Physiologie. Band 5, 1. 1883, S. 173 ff.
- Georg Löffler, Petro E. Petrides (Hrsg.): Biochemie und Pathobiochemie. 7. Auflage. Springer, Berlin u. a. 2002, ISBN 3-540-42295-1, S. 192.
- Renate Lüllmann-Rauch: Taschenlehrbuch Histologie. 4. Auflage. Thieme, 2012, ISBN 978-3-13-129244-5, S. 110 ff.
Einzelnachweise
- ↑ Paul Diepgen, Heinz Goerke: Aschoff/Diepgen/Goerke: Kurze Übersichtstabelle zur Geschichte der Medizin. 7., neubearbeitete Auflage. Springer, Berlin/Göttingen/Heidelberg 1960, S. 27 und 41.
- ↑ Hochspringen nach: a b H. Lodish, A. Berk, S. L. Zipursky u. a.: Molecular Cell Biology. W. H. Freeman, (englisch).
- ↑ Hochspringen nach: a b Tim Hunt: Molecular biology of the cell: the problems book. 6. Auflage. New York 2015, ISBN 978-0-8153-4453-7.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 10.05. 2026