Eukaryoten
| Eukaryoten | ||||
|---|---|---|---|---|

Verschiedene Eukaryoten | ||||
| Systematik | ||||
| ||||
| Wissenschaftlicher Name | ||||
| Eukaryota | ||||
| Chatton, 1925 |
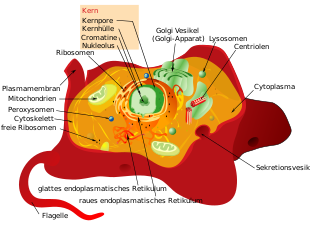
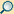
Eukaryoten oder Eukaryonten (Eukaryota) (von altgriechisch εὗ eu = ‚richtig‘, ‚gut‘ und κάρυον karyon = ‚Nuss‘, ‚Kern‘) sind eine Domäne der Lebewesen, deren Zellen (Eucyten) einen echten Kern und eine reiche Kompartimentierung haben (Tiere, Pflanzen und Pilze). Hierin unterscheiden sie sich von den beiden übrigen Domänen im Systematik der Lebewesen, den prokaryotischen Bakterien und Archaeen (letztere früher auch Urbakterien genannt), beide mit procytischen Zellen.
Merkmale
Die Zellen der Eukaryoten haben meistens einen Durchmesser von 10 bis 30 µm. Sie sind in der Regel wesentlich größer als die von Prokaryoten, ihr Volumen beträgt etwa das 100- bis 10.000-Fache. Für ein reibungsloses Funktionieren der zellulären Abläufe über größere Entfernungen innerhalb der Zelle sind ein höherer Organisationsgrad und eine Aufteilung des Zellraums in Kompartimente (abgegrenzte Räume) sowie Transport zwischen diesen Kompartimenten günstig. Aus diesem Grund sind eukaryotische Zellen mittels Zellorganellen strukturiert, welche wie die Organe eines Körpers verschiedene Funktionen ausüben. Das namensgebende Organell ist der Zellkern mit dem Hauptanteil des genetischen Materials der eukaryotischen Zellen. Weitere Gene kommen je nach Art in Mitochondrien (Organellen, die durch chemische Reaktionen Energie zur Verfügung stellen), in Einzelfällen auch in den ihnen verwandten Hydrogenosomen sowie in fast allen Plastiden (unter anderem in den Photosynthese betreibenden Chloroplasten) vor. Dem intrazellulären Transport dienen die Organellen des Endomembransystems.
Struktur und Form wird den eukaryotischen Zellen durch das Cytoskelett verliehen, das auch der Fortbewegung dient. Es ist aus Mikrotubuli, Intermediärfilamenten und Mikrofilamenten aufgebaut. Einige Eukaryoten, beispielsweise Pflanzen und Pilze, haben auch Zellwände, die die Zellen außerhalb der Cytoplasmamembran einschließen und ihre Form bestimmen.
Eine weitere Besonderheit der Eukaryoten liegt in der Proteinbiosynthese: Anders als Prokaryoten sind Eukaryoten in der Lage, mittels derselben DNA-Information durch alternatives Spleißen unterschiedliche Proteine herzustellen.
Mehrzelligkeit
Eukaryoten können Einzeller oder mehrzellige Lebewesen sein. Diese bestehen aus einer größeren Zahl von Zellen mit gemeinsamem Stoffwechsel, wobei spezielle Zelltypen bestimmte Aufgaben übernehmen. Die meisten bekannten Mehrzeller sind Eukaryoten, darunter die Pflanzen, Tiere und mehrzelligen Pilze.
Systematik
Der Name verweist auf das Vorhandensein eines Zellkerns (Eukaryota/Eukaryonta/Eucarya/Eukarya; zu altgriechisch εὖ eû ‚gut, echt‘ und κάρυον káryon ‚Nuss‘).
In der biologischen Systematik bilden die Eukaryoten eine der drei Domänen, also der höchsten Kategorien zur Klassifizierung von Lebewesen. Die aktuelle Systematik der Eukaryoten wurde von Adl et al. 2012 aufgestellt. Sie gliedert die Eukaryoten wie folgt:
- Amorphea
- super-group Amoebozoa, einzellige Organismen mit im Regelfall amöboider Gestalt, inkl.Schleimpilze und manche Amöben
- super-group Opisthokonta, inkl. Tiere und Pilze
- Diaphoretickes
- super-group Sar, inkl. Stramenopile (auch als Stramenopiles, Chromista oder Heterokonta bezeichnet)
- super-group Archaeplastida, inkl. Pflanzen
- super-group Excavata, einzellige Organismen, die mehrheitlich begeißelt sind
Daneben existieren zahlreiche Taxa mit unsicherer Stellung (incertae sedis), darunter unter anderem
- Cryptophyceae
- Centrohelida
- Haptophyta
- Telonemia
- Kamera lens, eine Geißeltierchenart und der einzige Vertreter der Gattung Kamera
- Picozoa
Entwicklungsgeschichte
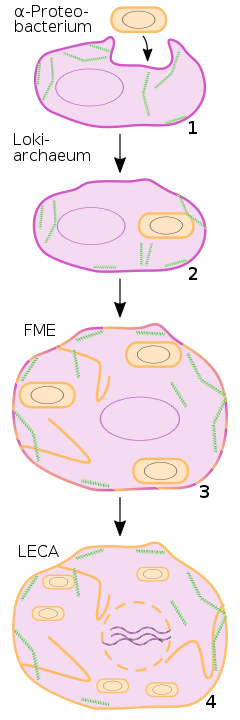
Die ältesten makroskopischen, mehrzelligen, möglicherweise eukaryotischen Fossilien sind 1,5 Milliarden Jahre alt. Es ist nicht bekannt, ob die hypothetischen evolutionären Vorläufer der Eukaryoten – die sogenannten Urkaryoten – bereits Organellen besaßen (diese wären aufgrund des nachteiligen Oberflächen-Volumen-Verhältnisses nötig gewesen) oder sie erst im Laufe ihrer Evolution hin zu den eigentlichen Eukaryoten erwarben. Die bekannteste Theorie zur Entstehung der Organellen ist die Endosymbiontentheorie, die besagt, dass Mitochondrien und Chloroplasten auf Bakterien zurückgehen, die von den zellbiologisch „primitiven“, räuberisch lebenden Urkaryoten ursprünglich als Nahrung aufgenommen und von denen einige durch Zufall nicht verdaut wurden, sondern eine Symbiose mit dem Urkaryoten eingingen.
Der letzte gemeinsame Vorfahr aller heutigen Eukaryoten – in der meist englischen Fachliteratur auch als Last Eukaryotic Common Ancestor, abgekürzt LECA, bezeichnet – sollte sowohl einen abgegrenzten Zellkern als auch Mitochondrien besessen haben, da alle bisher untersuchten Eukaryoten Mitochondrien, mitochondrienartige Organellen oder zumindest Kern-DNA von mitochondrialem Ursprung (übertragen durch lateralen Gentransfer) besitzen. Zwar führte Thomas Cavalier-Smith für rezente amitochondriale Einzeller mit Zellkern, die er für evolutionäre Relikte aus der Zeit vor LECA hielt, die Bezeichnung Archezoa ein, jedoch mehrten sich nachfolgend Hinweise darauf, dass es sich bei den Archezoen um sekundär amitochondriale echte Eukaryoten handelt, das heißt um Abkömmlinge von LECA.
Als Alternative zur Urkaryoten-Hypothese wird ein Erwerb von Mitochondrien durch Archaeen diskutiert (Wasserstoff-Hypothese) vor oder gleichzeitig mit dem Zellkern, der eventuell (ebenfalls) viralen Ursprungs (unter den NCLDVs) sein könnte („Viral eukaryogenesis“, ein Teil der „Out-of-Virus-Hypothese“). Als Kandidaten werden DNA-Viren wie die ‚Medusaviridae‘ gehandelt; und auch der ‚Pseudomonas Phage 201phi2-1‘ aus der Gattung Phikzvirus, Familie Myoviridae, ein Riesen-Bakteriophage, bildet bei der Infektion einer Bakterienzelle eine kernähnliche Struktur, die Proteine abhängig von ihrer Funktion trennt, diese kernartige Struktur und ihre Schlüsseleigenschaften wurden auch in den verwandten Phagen gefunden.
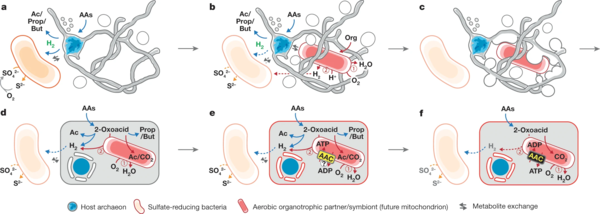
Nach 12 Jahren Forschungsarbeit berichteten Mikrobiologen 2019 erfolgreich ein, sich nur sehr langsam vermehrendes, Lokiarchaeon aus Tiefseeschlamm isoliert und kultiviert zu haben. Candidatus Prometheoarchaeum syntrophicum ist ein, nach Vergleich seines Genoms nahe mit den Eukaryoten verwandtes, extrem langsam wachsendes heterotrophes Archaeon, das Energie aus dem Abbau von Peptiden und Aminosäuren gewinnt und dabei Wasserstoff produziert und abgibt. Das Archaeon gedeiht nur in obligater Syntrophie mit Wasserstoff-metabolierenden Organismen, einer davon ein sulfatreduzierendes Delta-Proeobakterium der Desulfovibrionales. Die Autoren nehmen an, dass treibende Kraft der Endosymbiose, die zu den Eukaryoten führte, ein zweites, in ihrem Modellsystem nicht vorhandenes Bakterium, das künftige Mitochondrium, gewesen wäre, dessen Aufgabe zunächst primär die Beseitigung von für das obligat anaerobe Archaeon giftigen Sauerstoffs gewesen sei. Die Rolle des Mitochondriums im Energiestoffwechsel wäre demnach erst sekundär entstanden. Die Autoren halten eine Schlüsselrolle des Wasserstoff-Metabolismus selbst, wie in alternativen Szenarien der Endosymbiontentheorie vorgeschlagen, für unplausibel. Dem entsprechend wäre das Wasserstoff-verwertende Bakterium selbst nicht zum Endosymbionten geworden. Die Hypothese setzt einen frühen Ursprung des Mitochondriums in einer noch primitiven, Archaeen-artigen Wirtszelle voraus und unterscheidet sich darin von alternativen Szenarien wie der „Archezoa“-Hypothese von Thomas Cavalier-Smith, nach denen ein fast fertiger Pro-Eukaryot erst spät ein Mitochondrium erworben hätte. Dem entsprechend wäre ein „Einfangen“ des künftigen Mitochondriums über Phagocytose unplausibel, da es an den nötigen internen Strukturen wie einem Cytoskelett gefehlt hätte. Interessanterweise verfügt Prometheoarchaeum aber über tentakel-artige Zellfortsätze, mit denen ein ähnlicher Organismus im Archaikum den hypothetischen Endosymbionten alternativ quasi umschlungen haben könnte.
Die Entstehung der Chloroplasten der Photosynthese betreibenden Eukaryoten durch Endosymbiose stellt einen zeitlich späteren Vorgang dar. Zunächst wurden Cyanobakterien von nicht-phototrophen Eukaryoten aufgenommen, was möglicherweise in dieser Form (primäre Endosymbiose) nur einmal geschah. Die anderen Plastiden (Leukoplasten etc.) leiten sich von diesen ab.
Später entstanden komplexe Plastiden (mit mehr als doppelter Membran und ggf. Nukleosom) durch eine sekundäre Endosymbiose, d.h. (weitere) nicht-phototrophe Eukaryoten nahmen (aus der primären Endosymbiose entstandene) phototrophe Eukaryoten durch Endosymbiose auf (so etwa bei den Apicomplexa).
Forschungsgeschichte
Die Einteilung von Lebewesen in Prokaryoten und Eukaryoten wurde erstmals von Edouard Chatton für Protisten deutlich herausgestellt und 1925 veröffentlicht.
Diese Einteilung trat zunächst mit der Einführung des Drei-Domänen-Systems durch Carl Woese 1977 in den Hintergrund., ein Ansatz, der das zelluläre Leben in die drei Domänen Bakterien, Archaeen und Eukaryoten aufteilt.
Im Jahr 1984 wurde von James A. Lake und Kollegen die Eozyten-Hypothese vorgeschlagen: Man hatte entdeckt, dass bei der Form der Ribosomen in der Archaeen-Gruppe der Crenarchaeota (mit der ursprünglichen Bezeichnung Eozyten) und der Eukaryoten eine erstaunliche Ähnlichkeit besteht, dass aber die Form der Ribosomen sowohl bei den Bakterien als auch bei den Euryarchaeota, einer anderen Archaeen-Gruppe, deutlich davon abweicht. Es wurde daher vermutet, dass die Eukaryoten aus den Crenarchaeota hervorgegangen sind.
Trotz weiterer Hinweise in den 1980er Jahren bekam die Eozyten-Hypothese erst in den 2000er Jahren mit dem Fortschritt der Genom-Analyse neuen Auftrieb. In einer ganzen Reihe von Archaeen wurden Gene entdeckt, die man in ähnlicher Form auch in Eukaryoten findet. Entsprechend den Ergebnissen einiger Studien wurde in einer Abwandlung statt der Crenarchaeota die diesen nahestehende Gruppe der Thaumarchaeota als Ursprung der Eukaryoten vorgeschlagen. (Die Crenarchaeota und Thaumarchaeota werden zusammen mit einigen anderen Archaeen-Gruppen in der Supergruppe „TACK“ zusammengefasst).
Mit dem Aufkommen der Metagenomanalyse ließen sich seit 2015 in Proben aus der Nähe von Hydrothermalquellen Kandidaten für Archaeen-Gruppen identifizieren, die den Eukaryoten noch viel näher stehen müssen, als alle zuvor betrachteten Gruppen. Die erste Gruppe war die der „Lokiarchaeota“, gefunden bei einem hydrothermalen Vent Namens „Loki's Schloss“ im Arktischen Ozean> zwischen Mohns- und Knipovitsch-Kamm. Da diese Erkenntnisse lediglich aus einer Metagenom-Analyse stammen und man die betreffenden Mikroben noch bis auf weiteres nicht kultivieren kann, kommt allen fraglichen Taxa lediglich ein 'Kandidatenstatus' zu, gekennzeichnet durch die Anführungszeichen.
Die „Lokiarchaeota“ werden aufgrund von Ähnlichkeit im Genom mit einigen ebenfalls in diesem Zusammenhang jüngst vorgeschlagenen Gruppen („Thorarchaeota“, „Odinarchaeota“ und „Heimdallarchaeota“) zu einer Kandidaten-Gruppe „Asgard“ zusammengefasst, die unter den Archaeen den Eukaryoten somit am nächsten steht und ein Schwester-Taxon zur Supergruppe „TACK“ darstellt.
Um der engen Verwandtschaft der Archaeen mit den Eukaryonten Rechnung zu tragen, hat Thomas Cavalier-Smith sie bereits 2002 in ein gemeinsames Taxon Neomura gestellt, das damit eine Schwestergruppe zu den Bakterien darstellt.
Diese neue Einteilung wird unterstützt durch neuere Erkenntnisse, wonach die Verwendung des DNA-Genoms als Träger der Erbinformationen bei Bakterien einerseits und Archaeen und Eukaryonten andererseits unterschiedlichen (möglicherweise viralen: anderer Teil der „Out-of-Virus-Hypothese“) Ursprungs zu sein scheint. Der letzte gemeinsame Vorfahr aller heute bekannten Lebewesen wäre dann ein archaischer zellulärer Organismus der RNA-Welt (mit Ribosomen, d.h. Protein-Synthese) gewesen.
Weitere Unterstützung bekommt die Theorie durch Untersuchung des Aufbaus der membranständigen F- und V-/A-Typ-ATPasen. Die F-Typ-ATPasen der Mitochondrien und Chloroplasten sind mit denen der Bakterien homolog (wie nach der Endosymbiosetheorie zu erwarten). Die V-Typ-ATPasen an Zytoplasma-Membranen (Vakuolen) der Eukaryoten sind zu denen der Archaeen homolog, was eine Abstammung der Eukaryoten aus einem Zweig der Archaeen stützt. Vereinzelte Ausnahmen (F-Typ-ATPasen bei einigen Archaeen-Species und V-Typ-ATPasen bei einigen Bakteriengruppen) werden auf horizontalen Gentransfer zurückgeführt. Zentrale Untereinheiten der ATPasen sind über alle Lebewesen homolog, was für einen LUCA mit einer zumindest primitiven Membranhülle spricht.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 02.02. 2026